脂质纳米颗粒在当前SARS-CoV-2临床试验中的应用
1 BioNTech/辉瑞
BioNTech在SARS-COV-2试验的递送系统脂质成分主要是Acuitas的ALC-0315(表2)、DSPC、胆固醇和PEG-脂质。CureVac和伦敦帝国理工学院可能也使用ALC-0315或A9(表2)。BioNTech开始用四种mRNA编码的免疫原开发SARS-CoV-2疫苗,其中两种是核苷修饰的,一种是未修饰的,一种是自扩增的。
其中有关两种核苷修饰的mRNA:BNT162b1是一种较短的、约1 kb的序列,其编码刺突蛋白的受体结合结构域,由折叠三聚结构域修饰,通过多价显示增加其免疫原性;
另一种BNT162b2是较长的、约4.3 kb的薛烈,其编码二脯氨酸稳定的全长的膜结合刺突蛋白。BNT162b2最近获得了欧盟和美国的紧急批准。在一项临床前研究中,单次给药0.2、1和5μg的BNT162b2后,可检测到小鼠体内的结合抗体和中和效价,从**剂量到**剂量增加一个数量级,并在Th2细胞因子水平非常低的CD4+和CD8+脾细胞中引起强烈的抗原特异性Th1 IFNγ和IL-2反应。引流淋巴结也含有大量生发中心B细胞和CD4+和CD8+ T滤泡辅助细胞(Tfh)的数量也升高,这些细胞以前被认为是由mRNA LNP疫苗中LNP单独诱导。
在非人的灵长类动物中,30 μg或100 μg的免疫增强剂量引起的结合抗体和中和效价是人类恢复期组的10倍以上,并产生强烈的Th1偏向性T细胞反应,这对预防疫苗相关的增强型呼吸道疾病很重要。在6只的猕猴中,两次给药100 μg后在支气管肺泡灌洗液和鼻拭子中检测不到病毒效价。对较小的mRNA编码免疫原BNT162b1的1期临床试验中计划在第1天和第21天给药10、30和100 μg。中等剂量30 μg诱导的抗体结合和中和效价分别比人类恢复期组高30倍和3倍。由于第一次给药后出现严重的注射部位疼痛,因此未给予100 μg剂量的增强剂量。给药30 μg的增强剂量后100%的受试者产生轻度或中度的注射部位疼痛。第二次接种30 μg剂量后,几乎所有受试者都经产生了轻度或中度的全身不良反应,如发热、寒战或疲劳。该试验还证明了来自外周血单核细胞有强Th1偏向性T细胞反应。
一项2期临床试验比较了年轻(18-55岁)和年长(65-85岁)受试者组接种 BNT162b1和BNT162b2后的一些指标。老年组的结合和中和抗体效价略低,但仍高于恢复期组的受试者。与年轻组相比,老年组的不良反应严重程度也降低了。BNT162b2与BNT162b1相比,全身不良反应(发热、寒战、疲劳)的发生率**降低了约两倍。正是BNT162b2耐受性的增加推动了其3期临床试验,最近该试验宣布有效率达到94%,因为安慰剂组出现了162例COVID-19感ran,而接受两次30 μg剂量BNT162b2的接种组*发现8例感ran。
2 Moderna
在Moderna的研究中,核苷修饰的mRNA编码的免疫原是一种跨膜锚定的二脯氨酸稳定的预融合刺突,具有天然的呋喃裂解位点,并以LNP递送,该LNP参照MC3 LNP原型,但用脂质H (SM-102)替代MC3。mRNA LNP (mRNA-1273)接种剂量为1 μg而非0.1 μg,小鼠接种第1天和第21天,能诱导产生中和抗体。T细胞反应似乎是一种Th1/Th2平衡的反应,在小鼠的病毒模型中,两次给药1 μg(非0.1 μg)后,小鼠肺部和鼻甲的病毒效价降低到基线。以猕猴为实验对象,两次给药剂量为100 μg,给药后能产生高结合和中和效价以及在外周血中产生Th1偏向反应,同时也有强Tfh反应。两次给药10 μg剂量后效价和T细胞反应**降低。
同样,100 μg剂量能够将支气管肺泡灌洗液和鼻拭子中的病毒效价降低到基线水平,而10 μg剂量*在肺部有效。在一项1期临床研究中,每组15名患者,给药频率为间隔4周2次,给药剂量为25、100或250 μg,100 μg剂量的结合和中和效价比恢复期高约10倍,相当于25 μg的恢复期。所有受试者在100 μg和250 μg剂量下均产生了不良反应,250 μg组的14名受试者中有3名发生了严重的不良反应,并被停药。在随后对老年患者(56-71岁和71岁以上)进行的1期临床研究中,发现25 μg和100 μg剂量产生的结合抗体效价高于恢复期血浆中的,而中和效价与100 μg相当,但低于恢复期的25 μg。
即使是在老年组也有约80%的患者在第二次接种疫苗后仍出现不良反应。外周血分析显示CD4 T细胞反应是Th1偏向型的。与25 μg剂量相比,100 μg剂量的中和效价更高,因此进一步进行了3期临床试验,中期结果显示,安慰剂组出现了90例COVID-19感ran,而接种疫苗组只有5例感ran,防护有效率达到94.5%。一**委员会对Moderna的3期临床试验的中期分析结果表明,出现严重不良反应包括:9.7%的接种者出现疲劳、8.9%的接种者出现肌肉疼痛、5.2%的接种者出现关节tong、4.5%的接种者出现**,而在辉瑞/BioNTech的3期临床试验中,出现疲劳的频率较低,只有3.8%,**为2%。
3 CureVac
CureVac mRNA LNP (CVnCoV)采用的是一种非化学修饰的、序列工程mRNA,它编码一种二脯氨酸稳定的全长的S蛋白,采用Acuitas LNP递送技术,可能使用可电离脂质ALC-0315。小鼠实验中剂量为2 μg时,对两次给药之间的周数(从1到4不等)进行了研究,发现较长的时间间隔在Balb/c小鼠中产生更高的效价和T细胞反应以及平衡的Th1/Th2反应。产生中和抗体需要第二次给药,两次给药0.25 μg不足以产生中和抗体。在叙利亚金黄地鼠实验中,两次给药10 μg (非2 μg)能够将肺(非鼻甲)中的病毒效价降低到基线。在剂量为2-12 μg的1期临床试验中,*在**剂量12 μg时发现中和效价达到恢复期血清水平,这使得较高剂量(16和20 μg)被纳入正在进行的2期临床试验。所有给药剂量为12 μg的患者在每次给药后都产生了全身不良反应,大多数为中度和重度,而> 80%的患者在局部注射部位产生了轻度和中度疼痛。
4 TranslateBio
Translate Bio使用的是一种非修饰的mRNA,其编码双突变的二脯氨酸稳定的刺突蛋白,采用LNP技术平台,使用可电离脂质C12-200,C12-200很可能是近期的基于ICE-或半胱氨酸的可电离脂质家族合成的候选物。在Balb/c小鼠实验中,发现在0.2-10 μg范围内两次给药使结合和中和效价远高于恢复期水平。在非人的灵长类动物实验中,15、45和135 μg剂量均产生超过人类恢复期的效价,并且其免疫反应也是Th1偏向型的。
5 Arcturus
Arcturus使用的是一种自扩增的、全长的、未修饰的mRNA,其编码融合前SARS-CoV-2全长的刺突蛋白,该LNP中使用带有硫酯的可电离脂质,通过两个额外的酯基将含胺的头部和脂质尾部相连接。这个脂质家族中两种可能的可电离脂质是脂质10a(在[111]的表4中)或脂质2,2 (8,8) 4CCH3(在[57]的第33页上)(表2)。
后者有三个分支,类似于Moderna脂质H,但有一个可降解的硫酯连接到头部。自扩增mRNA的一个特征是荧光素酶基因的表达在肌肉注射给药一周后保持在相当恒定的水平,而常规mRNA的表达则迅速下降。在C57BL/6小鼠中,单独接种疫苗使体重减轻和临床评分增加。在具有高水平抗原特异性T细胞反应的Th1偏向反应中,只需要对小鼠以2 μg或10 μg(非0.2 μg)的剂量单次给药,就可以使中和效价达到100以上。在K18-hACE2小鼠致死模型中,单次给药2 μg或10 μg也可以100%保护小鼠,并且小鼠体重没有减轻,且肺和脑的病毒效价降低到基线。Arcturus已经完成了接种剂量为1-10 μg的1期临床试验,并选择使用7.5 μg作为接种剂量进行3期临床试验。
6伦敦帝国理工学院
伦敦帝国理工学院使用了一种由Acuitas LNP技术递送的、自扩增的编码预融合的稳定的刺突蛋白的mRNA,该蛋白在由脂质A9的**中有所描述(表2)。在Balb/c小鼠中两次注射0.01 μg至10 μg的剂量后,产生了非常高的剂量依赖性抗体和中和效价。该反应是强Th1偏向型,与较低的0.1和0.01 μg剂量相比,10和1 μg的剂量产生了高三倍的抗原特异性脾细胞反应。该疫苗即将开始1期临床试验。
7 朱拉隆功大学,宾夕法尼亚大学
朱拉隆功大学与宾夕法尼亚大学合作,使用Genevant LNP技术开发一种天然刺突免疫原核苷修饰的mRNA LNP,采用的脂质可能是CL1。他们的目标是于2021年第一季度开始第一阶段临床试验,并于2021年第四季度开始向泰国和七个周边中低收入国家供应疫苗。
8 Providence Therapeutics
Providence Therapeutics获得了加拿大卫生部的授权通知,可以对PTX-COVID-19B mRNA LNP疫苗进行人体临床试验。对编码受体结合域(具有或不具有呋喃裂解位点突变的全长刺突蛋白)的三种候选mRNA进行临床前研究,按照免疫增强方法以C57BL6小鼠为实验对象以20 μg的剂量给药。
该疫苗使用来自Genevant的未公开脂质(可能与表2中的CL1相似)的临床前数据,显示出全长的和呋喃突变的有效载荷具有强大的中和效价。第一阶段临床试验计划于2021年第一季度开始,同时疫苗的生产和销售计划于同年获得监管机构的批准。
9 储存和供应
实验室制造的大多数RNA LNP可以在4℃下保存几天,但随后出现颗粒尺寸增加和生物活性逐渐丧失(如荧光素酶表达)的特点。在以前的siRNA LNP方中,通常由于LNP聚集导致尺寸随着时间增大。为了mRNA LNP疫苗能稳定的储存和供应,需要采用冷冻形式。Moderna的COVID-19疫苗需要在-25℃至-15℃之间储存,在2℃至8℃之间可稳定储存30天,在8℃至25℃可稳定储存12小时。
辉瑞/BioNTech的COVID-19疫苗需要在-80℃至-60℃之间储存,解冻之后在2℃至8℃之间储存最多5天,然后在注射前用盐水稀释。在储存和运输过程中,辉瑞疫苗所需的极低温度比Moderna疫苗所需的常规冷冻温度更难达到。这些温度差异背后的原因并不明显,因为两种疫苗都含有类似的高浓度蔗糖作为冷冻保护剂。Moderna的mRNA LNPs被冷冻在Tris和醋酸盐两种缓冲液中,而辉瑞/BioNTech疫苗*使用磷酸盐缓冲液。众所周知,磷酸盐缓冲液不适合冷冻,因为它们容易沉淀而且结晶会引起pH突变。冻干工艺对mRNA LNPs来说是一个技术难点。然而,Arcturus已经声明,他们的COVID-19 mRNA疫苗在冻干形式中是稳定的,尽管这种冻干制剂的温度稳定性尚未公开,但这可能会**简化供应。
脂质纳米颗粒
许多脂质样实体,称为脂质类化合物类脂,最初是为siRNA递送而开发的,随后用于mRNA递送。C12-200就是一个例子(表2),由于其通过静脉给药在肝细胞基因沉默中的高效性,而在类脂家族中脱颖而出。为了实现高效的肝脏靶向基因沉默,C12-200与MC3 Onpattro原型相同的脂质相结合,即50%可电离脂质、10% DSPC、38.5%胆固醇和1.5% PEG-脂质。
后来的一项研究发现,通过将可电离脂质的百分比降低到35%,同时将可电离脂质与核酸的重量比从5增加到10,并用促细胞融合的不饱和DOPE取代DSPC,可以将C12-200对同一肝脏靶点的mRNA递送效率提高7倍。
有趣的是,这种优化的方将mRNA表达提高了7倍,但没有改变siRNA的沉默效率。在这种方中,C12-200也被研究用于小鼠和非人灵长类动物的mRNA介导的蛋白质替代疗法,但是当皮下注射时,通过组织学观察,它会产生强烈的炎症反应。C12-200是一种小分子树状聚合物,具有五个烷基链和五个氮原子,根据ACDLabs Percepta等商用软件进行电离分析,发现其中三个似乎是可质子化的(表2)。另一种树枝状大分子脂质,5A2-SC8,在观察过程中发现对肝脏具有高siRNA传递效率,并且还具有五个氮原子和五个短烷基链(表2)。
类脂5A2-SC8对于mRNA递送的效率很低,除非通过将可电离的脂质摩尔分数降低到24%,使用DOPE代替DSPC,并增加其他脂质比例来改变其方参数,但是同时5A2-SC8与mRNA的重量比将增加到20。
这些方的改变似乎是这些树枝状大分子型类脂成为有效的mRNA递送载体所必需的,这可能是因为它们具有多质子的头部和树枝状大分子结构。另一种非常高分子量的修饰树枝状大分子被用于递送编码流感、埃博拉和弓形虫免疫原的自扩增mRNA,并且在小鼠实验中,在单次给药40 μg的高剂量或免疫增强方法注射4 μg后(这对复制RNA也是高剂量),发现可以避免三种病原体的感ran。最近对类脂进行研究发现,与其他类脂相比,这种小的三氮树枝状大分子的四个烷基链末端增加一个碳支链,肝脏表达能力提高了10倍以上。
这种效价的增加与LNP的pKa值没有相关性,但与pH为 5时TNS染料的**荧光有相关性,这表明内涵体质子化的幅度与mRNA表达相关,推测可能是促进内涵体逃逸。根据分子形状假说,增加的碳支链也可以产生一个更锥形的结构,从而产生更多的膜破裂。
编码抗体基因的mRNA LNP的递送
目前市场上有70多种单克隆抗体(mAbs),全球销售额为1250亿美元。使用mRNA编码的抗体具有以下优势,包括有益于天然翻译后修饰的内源性蛋白质合成,以及是一种不需要细胞培养和对蛋白质产品大量纯化和表征的简化生产方法。通过将编码VRC01(一种针对HIV-1的中和抗体)轻链和重链的纯化核苷修饰的mRNA包封到Acuitas LNPs中,显示出递送mRNA编码的mAbs可能用于被动免疫。
以 Balb/c小鼠为实验对象,静脉注射30 μg mRNA LNP(靶向肝细胞),表达mAbs超过一周,血清水平达到150 μg/ml,高于直接注射600 μg mAbs,每周注射能够保持血清水平在40 μg/ml以上。CD34-NSG人源化小鼠注射30 μg和15 μg的mRNA LNP可以抵御24小时后的HIV-1攻击,入侵2周后的血清病毒RNA复制分析也证明这一观点。
CureVac的一项研究证实了**性非修饰mRNA编码抗体的可行性,该研究也使用了Acuitas LNPs,其中选择了对多种狂犬病毒株具有**中和能力的IgG mAbs,以及针对肉毒杆菌**的纯重链Vh结构域(VHH)中和剂。还生产了一种靶向CD20的、mRNA编码的利妥昔单抗,其中CD20是非霍奇金淋巴瘤**的**。
动物实验通过静脉注射使用的是靶向肝细胞的Acuitas LNP。小鼠单次给药40 μg产生的血清抗体水平刚刚超过10 μg/ml,1个月后逐渐下降到1 μg/ml。相同剂量下,VHH单结构域中和剂产生的抗体水平高10倍,但由于缺乏Fc区,半衰期*有几天。当在狂犬病病毒的致命攻击之前1天或之后2小时,对小鼠单次静脉注射40 μg也能够完全保护小鼠。
同样,在致命的肉毒杆菌**攻击后6小时单次给药40 μg也完全保护了动物。第三个攻击模型是将Raji-luc2 B细胞淋巴瘤细胞静脉移植并且生长4天,然后在18天内,在Acuitas LNP中5次给药10或50 μg mRNA编码的利妥昔单抗,结果是保护了所有动物,并且50 μg剂量能够完全消除**生长。
将T细胞募集到**细胞的双特异性抗体也被编码在修饰的mRNA结构中,并使用商业转染试剂TransIT在体内递送,该试剂在肝脏递送方面不如目前的LNP有效。
该mRNA结构可维持循环和生物活性的双特异性抗体超过6天,而相同5 μg剂量的蛋白质-双特异性抗体在**后减少至接近基线。第二项研究也是使用VHH形式的双特异性抗体进行的,其中一个结合保守的甲型流感基质蛋白2外域(M2e)的VHH基因与另一个小鼠Fcγ受体IV (FcγRIV) 特异性结合的VHH基因相连,以便将表达FcγRIV的先天免疫细胞募集到表达M2e的流感感ran细胞中。
这些核苷修饰过结构的mRNA使用DOTAP/胆固醇制备的LNPs递送,通过气管内滴注到小鼠肺中,4小时后,用致死剂量的流感病毒攻击,80%的小鼠免受致死剂量的影响,尽管它们产生明显的体重减轻,并且DOTAP/胆固醇mRNA纳米粒导致粒细胞暂时流入肺部,同时血清IL-6细胞因子水平也有所升高。
**,在基孔肯雅感ran幸存者的B细胞中发现的一种有效中和抗体被编码在一种核苷修饰的mRNA结构中,该结构由一个可能含有MC3或脂质5的LNP递送。静脉注射0.5 mg/kg(10 μg)编码mAb的mRNA,24小时预注射的小鼠获得了抗病毒攻击的保护作用,而注射蛋白mAb需要2 mg/kg的剂量。在感ran后4小时,对小鼠注射高剂量10 mg/kg (200 μg),使小鼠避免感ran。非人的灵长类动物研究中发现,3 mg/kg(9 mg)的高剂量产生的短暂毒性(包括脾脏增大和CCL2血清水平升高)最小,且注射后几个月仍可检测到抗体。基于这些结果,Moderna启动了一项1期临床试验,并公布了阳性结果,其中0.1和0.3 mg/kg的剂量耐受性良好,并且mAb的血清水平在1–14 μg/mL的范围内,预计单次给药后可对基孔肯雅病毒具有长达16周的免疫作用。
脂质纳米颗粒组装和结构
目前mRNA脂质纳米颗粒的生产方法是利用微流体或T型接头,把含有疏水性质的乙醇相和含有mRNA的水相在pH为4的缓冲液(如乙酸)中混合(图2)。现有的方法(如薄膜蒸发法和乙醇注入法),由于纳米粒子粒径不稳定、mRNA包封率较低、难以扩大规模而很少使用。微流体混合的优点是能够将乙醇中非常小体积的脂质与几十μL水溶液中的mRNA混合,从而可以筛选许多成分和方参数。另一方面,T型接头混合器是大批量商业生产mRNA LNPs的通用方法,例如目前临床试验中使用的方法。
最近的一份期刊表明,这两种方法都可以生产有类似大小和形态的LNP。两种溶液的快速混合是控制粒径< 100 nm的关键,从而避免了其他生产方法所需的尺寸减小的需要(挤出、超声处理)。如图2所示,由这些溶液组装和形成LNP的过程是由疏水力和静电力驱动的。四种脂质(可电离脂质、DSPC、胆固醇、PEG-脂质)最初可溶于乙醇,可电离脂质非质子化且呈电中性(图2A)。通常将一体积的含脂质的乙醇溶液与三体积的mRNA在pH=4的醋酸盐缓冲液中混合,这使得脂质接触缓冲液时,它们变得不溶于3∶1的水/乙醇溶剂,并且可电离的脂质质子化携带正电荷,然后与mRNA的带负电荷的磷酸骨架静电结合(图2B),形成包封mRNA的脂质颗粒,同时脂质在主要是水的悬浮液中变得难溶。
这一过程中的一个关键成分是PEG-脂质,因为PEG链是亲水性的,从而包覆颗粒,并决定其最终的热力学稳定尺寸大小。通过改变PEG的摩尔分数,可以控制LNP的大小,例如,LNP颗粒大小为100 nm时PEG-脂质的摩尔分数为0.5%,而43 nm时PEG-脂质的摩尔分数为3%。最近一个研究显示,当mRNA LNP悬浮液在水溶液缓冲液中稀释或透析以提高pH并去除乙醇时,LNP的结构和大小在混合后继续发生变化。水相和脂质相混合时初始pH值接近5.5,使可电离脂质质子化,其LNP的pKa接近6.5,可以与mRNA结合和包裹(图2B,C)。
之后通过稀释、透析或切向流过滤提高pH可以中和可电离脂质,直到pH为7.4时可电离脂质基本不带电(图2D)。当可电离脂质变为电中性时,它也变得更难溶解,导致形成更大的疏水类脂结构域,从而促进LNP的融合,使LNP的尺寸增大,LNP的中心形成无定形的电子致密相,主要包含与mRNA结合的可电离脂质。据估计,在这个过程中,多达36个囊泡可以融合形成最终的LNP(图2C,D)。使用FRET对证实了这一融合过程,并进一步看到了PEG-脂质在这一过程中的作用,因为混合后加入PEG-脂质与混合前加入PEG-脂质以相同的方式控制最终LNP的大小。这项研究和另一项使用中子散射方法的研究也表明,DSPC在LNP的**PEG层的下面形成了一个双层结构,其**主要是与mRNA结合的可电离脂质(图2D),同时认为胆固醇分布在整个LNP中。
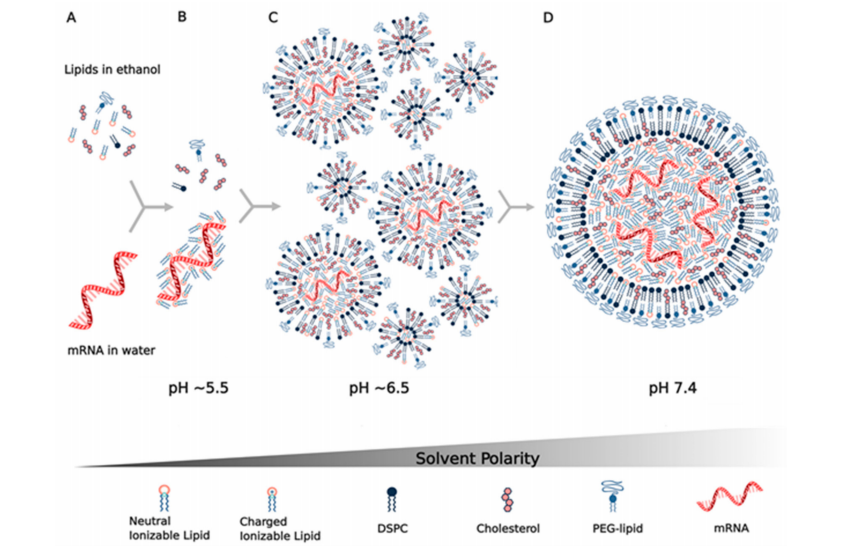
图2:mRNA脂质纳米颗粒的组装是通过(A)在微流体或T型接头混合器中将四种脂质(可电离脂质、DSPC、胆固醇、PEG-脂质)在乙醇中与mRNA在pH4左右的水溶液缓冲液中快速混合而实现的。
(B)当可电离脂质与水相接触时,在pH~5.5范围内质子化,该pH介于缓冲液的pKa和可电离脂质的pKa之间。
(C)可电离脂质与mRNA的阴离子磷酸骨架静电结合,同时它在水相中经历具有疏水性,驱动囊泡的形成和mRNA的包裹。
(D)在初始囊泡形成后,通过稀释、透析或过滤提高pH,中和可电离脂质,使其疏水性增强,从而驱动囊泡融合,导致可电离脂质与mRNA进一步包裹在脂质纳米粒的内部。通过给LNP提供亲水性的外表、确定其热力学稳定的尺寸大小,PEG-脂质的含量停止融合,在PEG-脂质层的下面形成DSPC的双层结构。
本文文献的参考文献格式:
Michael D. B;Manuel J. C;Suman A;Mikell P;Mohamad G A;Drew W. Nanomaterial Delivery Systems for mRNA Vaccines. Vaccines 2021, 9, 65
参考文献:
1. Zimmer, C.; Corum, J.; Wee, S.-L. Coronavirus Vaccine Tracker. In The New York Times; The New York Times Company: New York,NY, USA, 2020.
2. Moderna’s COVID-19 Vaccine Candidate Meets its Primary Efficacy Endpoint in the First Interim Analysis of the Phase 3 COVE Study|Moderna, Inc. Available online: https://investors.modernatx.com/news-releases/news-release-details/modernas-covid-19-vaccine-candidate-meets-its-primary-efficacy/ (accessed on 29 November 2020).
3. Pfizer and BioNTech Conclude Phase 3 Study of COVID-19 Vaccine Candidate, Meeting All Primary Efficacy Endpoints|BioNTech.Available online: https://investors.biontech.de/news-releases/news-release-details/pfizer-and-biontech-conclude-phase-3-study-covid-19-vaccine/ (accessed on 29 November 2020).
4. Gómez-Aguado, I.; Rodríguez-Castejón, J.; Vicente-Pascual, M.; Rodríguez-Gascón, A.; Solinís, M.á.; del Pozo-Rodríguez, A.Nanomedicines to Deliver mRNA: State of the Art and Future Perspectives. Nanomaterials 2020, 10, 364. [CrossRef]
5. Hajj, K.A.; Whitehead, K.A. Tools for Translation: Non-Viral Materials for Therapeutic mRNA Delivery. Nat. Rev. Mater. 2017,2, 17056. [CrossRef]
6. Kowalski, P.S.; Rudra, A.; Miao, L.; Anderson, D.G. Delivering the Messenger: Advances in Technologies for Therapeutic mRNA Delivery. Mol. Ther. 2019, 27, 710–728. [CrossRef]
7. Li, B.; Zhang, X.; Dong, Y. Nanoscale platforms for messenger RNA delivery. Wiley Interdiscip.Rev.Nanomed. Nanobiotechnol.2018, e1530. [CrossRef]
8. Van Hoecke, L.; Roose, K. How mRNA Therapeutics Are Entering the Monoclonal Antibody Field. J. Transl. Med. 2019, 17, 54.[CrossRef]
9. Wadhwa, A.; Aljabbari, A.; Lokras, A.; Foged, C.; Thakur, A. Opportunities and Challenges in the Delivery of mRNA-Based Vaccines. Pharmaceutics 2020, 12, 102. [CrossRef]
10. Wahane, A.;Waghmode, A.; Kapphahn, A.; Dhuri, K.; Gupta, A.; Bahal, R. Role of Lipid-Based and Polymer-Based Non-Viral Vectors in Nucleic Acid Delivery for Next-Generation Gene Therapy. Molecules 2020, 25, 2866. [CrossRef]